
RbAp48: The Memory Gene

2013, a group of researchers from various departments of Columbia University led a study regarding memory loss related to aging. The study aimed in particular to look for molecular patterns that could mark this phenomenon and, since the difference was already stressed on an anatomical level, tried to highlight the difference between age-related memory loss and Alzheimer disease (AD) on a molecular level. As a result, the researchers identified the gene RbAp48 as a possible responsible for age-related memory loss. The hippocampus was proven to be involved in the formation of long-term memories and, therefore, its role in pathologies whose result was the loss of such memories was investigated: in previous studies, researchers have proven that there was a specific anatomical pattern for each disease. More specifically, the entorhinal cortex (EC) and the hippocampus’ subregion CA1 were involved in AD while, simple age-related memory, loss was correlated to both the dentate gyrus (DG) and the subregion CA3 [1-4]. The researchers from Columbia University started their study by analysing healthy human brain’s DG and EC post mortem tissue with subjects ranging from the age of 33 to the age of 89 [10]. The two tests ran (micro array assay and protein level assay) suggested that the protein RbAp48 was the one whose deficiency best correlated with age, hence highly varying in the DGs, and whose concentration was unaffected in the ECs making this protein irrelevant for AD [5, 6]. Moreover, RbAp48 was already listed among other proteins vital to the correct transcriptional regulation in brain, to the cAMP-PKA-CREB pathway and to histones’ acetylation: all these mechanisms were known to be required for normal hippocampal function and aging in the mouse [7].
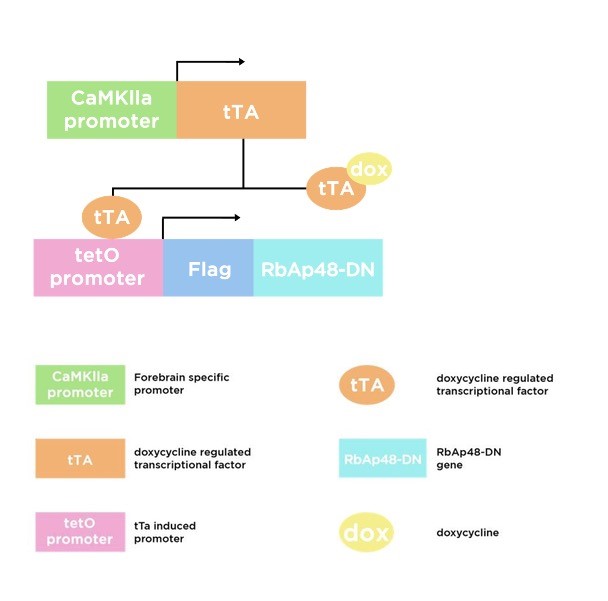
Figure 1: tTa vector structure
Since the impairment of this protein in a KO animal model might result in developmental issues, the study had to feature a particular animal model that allowed to study the loss of RbAp48 from a specific age and in a specific brain area and also the subsequent protein recovery: the researchers generated a double-transgenic (DT) mice that expressed a dominant-negative form of RbAp48 (RbAp48-DN) which was lacking the first 54 N-terminal amino acid residues, vital for the interaction with histone H4, and which was modulated by the administration of doxycycline (Figure 1).Data found in this animal model were consistent with the ones found in human brains indicating a high level of RbAp48 in DGs of young mice and a significantly lower concentration in older mice’s DG [11]. To exclude that anxiety could impair the cognitive tests’ results, both WT and DT mice underwent the elevated plus maze and the open field test. WTs and DTs resulted having similar level of anxiety. It was then investigated whether inhibiting RbAp48 in the young adult mouse (3,5 month old) affected memory. By using a novel object recognition test and the Morris water maze the researchers confronted young adult DT mice with the performances of old WT mice and, not only was it proven that RbAp48 was responsible for memory loss, but also that the recovery of the protein (by administration of doxycycline) meant the recovery of the memorization function. To furtherly strengthen the aforementioned data, the researchers also tried to upregulate the protein in aged mice by introducing a lentiviral gene transfer which lead to doubling the concentration of RbAp48. The results showed that a higher concentration of the protein significantly improved memory performance in both the novel object recognition and Morris water maze tasks and that this improvement was independent from neurogenesis. Eventually, this study investigated the effect of RbAp48-DN expression on histone acetylation in the DG and on HAT activity of CBP (a critical mechanism for memory). It was found that inhibition of RbAp48 caused a reduction in acetylation of both H4 and H2B which proved to be reversible if the protein was restored to normal and that the CBP HAT activity, but not the amount of CBP protein, was significantly lower in the DG of RbAp48-DN–expressing mice than in controls [8, 9].
The researchers conclude that altogether the data suggest that RbAp48 plays an important role in hippocampus-related memory processes and that it has a causal role in cognitive aging. Although the article is technically sound and provides a lot of new information, it leaves more than one question open about the possibility to apply it on human beings as well. It would be of utmost interest to understand whether the level of schooling and or brain activity (intended as reading frequently, solving riddles, writing etc) of the subject may affect the ability to produce memories. It must also be taken into account that nowadays a person is subject to incredible stresses and more often than not the life style led is not at all optimal. Therefore, a question that needs to be answered is whether bad habits such alcohol consumption, smoking and a sedentary life style affect or not the activity of RbAp48 and whether, before administrating any sort of drug, it is possible to prevent this protein loss by ameliorating our life styles.
References
- Small SA, Schobel SA, Buxton RB, Witter MP, Barnes CA. A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat Rev Neurosci. 2011;12:585–601
- Braak H, Alafuzoff I, Arzberger T, Kretzschmar H, Del Tredici K. Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol. 2006;112:389–404.
- Gómez-Isla T, Price JL, McKeel DW, Jr, Morris JC, Growdon JH, Hyman BT. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. J Neurosci. 1996;16:4491–4500
- Small SA, Perera GM, DeLaPaz R, Mayeux R, Stern Y. Differential regional dysfunction of the hippocampal formation among elderly with memory decline and Alzheimer’s disease. Ann Neurol. 1999;45:466–472
- Yassa MA, Lacy JW, Stark SM, Albert MS, Gallagher M, Stark CE. Pattern separation deficits associated with increased hippocampal CA3 and dentate gyrus activity in nondemented older adults. Hippocampus. 2011;21:968–979
- Yassa MA, Lacy JW, Stark SM, Albert MS, Gallagher M, Stark CE. Pattern separation deficits associated with increased hippocampal CA3 and dentate gyrus activity in nondemented older adults. Hippocampus. 2011;21:968–979
- Alarcón JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: A model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration.Neuron. 2004; 42:947–959. [PubMed: 15207239]
- Zhang Q, Vo N, Goodman RH. Histone binding protein RbAp48 interacts with a complex of CREB binding protein and phosphorylated CREB. Mol Cell Biol. 2000; 20:4970–4978. [PubMed: 10866654]
- Korzus E, Rosenfeld MG, Mayford M. CBP histone acetyltransferase activity is a critical component of memory consolidation. Neuron. 2004; 42:961–972. [PubMed: 15207240]
- Lewandowski NM, Small SA. Brain microarray: Finding needles in molecular haystacks. J Neurosci. 2005; 25:10341–10346. [PubMed: 16280569]
- Pavlopoulos E, Jones S, Kosmidis S, Close M, Kim C, Kovalerchik O, Small SA, Kandel ER (2013) Molecular mechanism for age-related memory loss: the histone-binding protein RbAp48. Sci Transl Med 5(200):200ra115.